
Un entraînement intense active la synthèse des protéines, mais seulement si la bonne nutrition est là pour la soutenir. Si vous lisez l'article que John Meadows et moi avons écrit intitulé Maximize Protein Synthesis, ou si vous avez lu quelque chose que T Nation dit sur le sujet depuis des années, vous connaissez probablement le concept de «fenêtre anabolique» et son importance de la nutrition péri-entraînement.
Ce qui se passe au niveau cellulaire dans les heures qui suivent l'entraînement est prédictif des gains à long terme. Profitez de cette «fenêtre anabolique» et vous grandirez comme jamais auparavant. Manquez-le constamment, et bonne chance.
Obtenir les bons macronutriments au bon moment est la clé, mais les macros ne sont qu'une partie de la vue d'ensemble. Un aspect important mais souvent négligé de la synthèse des protéines musculaires est le volume cellulaire. Le volume cellulaire n'est pas seulement cosmétique; c'est le principal moteur du transport des acides aminés, travaillant également en coulisse pour activer la synthèse des protéines et supprimer la dégradation des protéines.
Un muscle plein / volumisé est un muscle anabolique. Alors que nous savons depuis plus de 20 ans que le gonflement des cellules inhibe la dégradation des protéines et stimule la synthèse des protéines dans certaines cellules (1-3), jusqu'à récemment, le mécanisme liant le volume cellulaire à la synthèse des protéines était un mystère.
Ce que nous savons maintenant, c'est que la synthèse des protéines est contrôlée par l'enzyme mTOR, qui est activée par le stress mécanique, les facteurs de croissance et la leucine.
Bien que ces trois éléments soient importants pour le stimulus d'entraînement, la signalisation mTOR dépend également du volume cellulaire.(4) Ceci est particulièrement important dans le muscle squelettique, où la volumisation cellulaire active la synthèse du glycogène et des protéines et inhibe la dégradation des protéines.(5, 6)
La percée scientifique menant au lien entre le volume cellulaire et la synthèse des protéines s'est produite en 2005, lorsqu'un groupe de scientifiques a découvert qu'il fallait plus que de la leucine pour activer le mTOR - la glutamine est également nécessaire.(7)
Cela a été une surprise. Bien que la glutamine soit considérée comme un acide aminé «conditionnellement essentiel» qui limite la dégradation des protéines lors de traumatismes graves ou de stress, elle n’a jamais été liée à l’activation de mTOR.
La glutamine était nécessaire pour l'absorption de la leucine et la volumisation cellulaire, qui sont toutes deux nécessaires pour activer la synthèse des protéines. Les auteurs ont ensuite montré que la glutamine cellulaire épuisement entraîne non seulement une réduction du volume cellulaire, mais réduit également la capacité de la leucine à activer la synthèse des protéines.(7)
Cette découverte était énorme, car elle fournissait un lien direct entre la glutamine, la volumisation cellulaire et la synthèse des protéines. Pour la première fois, il a été montré que la glutamine est nécessaire à l'activation de la synthèse protéique par la leucine.
Alors que cette étude a suggéré que la glutamine est une pièce très importante du puzzle liant le volume cellulaire et la synthèse des protéines, le mécanisme exact n'a été mis au point qu'en 2009, lorsque Nicklin et al. découvert que l'exportation de glutamine est couplée à l'importation de leucine et à l'activation de mTOR.(8)
Pour faire entrer la leucine dans la cellule, il y a une période initiale de «chargement de glutamine.«Cela aspire également l'eau, ce qui augmente le volume de la cellule. Après la phase de «chargement de glutamine», la glutamine est exportée hors de la cellule en échange de l'importation de leucine.
Nicklin et coll. ont également découvert que les niveaux cellulaires de glutamine limitent le taux d'activation de la synthèse des protéines par la leucine. Lorsque les cellules ont été traitées simultanément avec de la glutamine et un mélange EAA contenant de la leucine, l'activation de la synthèse des protéines a été retardée de 60 minutes. Lorsque ces mêmes cellules étaient «préchargées» en glutamine, la synthèse des protéines était activée dans les 1 à 2 minutes suivant l'administration de leucine aux cellules.
Ce résultat était important car il expliquait le temps de latence pour l'activation de la synthèse protéique par la leucine dans ce modèle expérimental.
Ces résultats jettent enfin un peu de lumière sur la machinerie cellulaire qui régule le transport des acides aminés, et comment elle est couplée au contrôle de la synthèse des protéines.
Ce travail doit être interprété avec une certaine prudence, cependant. Une mise en garde importante à ces études était qu'elles ont été réalisées in vitro (i.e. dans les cultures cellulaires) où la régulation ou la synthèse protéique est beaucoup plus simple. Les cellules musculaires sont capables de fabriquer de la glutamine selon les besoins à partir d'autres acides aminés, et «l'épuisement de la glutamine» dans ce modèle de culture cellulaire n'est pas représentatif de situations plus physiologiques in vivo.
Des taux élevés de synthèse des protéines ne peuvent pas être maintenus indéfiniment dans le tissu musculaire avec ou sans supplémentation en glutamine. La glutamine peut cependant être utilisée pour soutenir stratégiquement la synthèse des protéines en optimisant la volumisation cellulaire pendant la période post-entraînement.

Les cellules sont très occupées et il existe de nombreux canaux ioniques liés à la membrane et des protéines de transport qui régulent le trafic entrant et sortant de la cellule. Il existe deux classes de transporteurs d'acides aminés en particulier qui sont importants ici: les transporteurs d'acides aminés «System L» et «System A» sont les plus étroitement liés à la signalisation mTOR et à la synthèse des protéines.(8-10)
L'activité des transporteurs du Système A et du Système L est couplée, ce qui permet à la leucine et aux autres BCAA d'être absorbés dans la cellule.(11) Les transporteurs du système L sont responsables de l'afflux de leucine et des autres BCAA en échange de l'efflux de glutamine.
Les transporteurs du système A fonctionnent selon un mécanisme différent, cependant, où la glutamine est couplée à l'absorption de sodium.(12, 13) Le couplage entre l'absorption de sodium et les transporteurs d'acides aminés du Système L / Système A est appelé Transport Actif Tertiaire (TAT en abrégé). C'est TAT qui conduit finalement la leucine à l'intérieur de la cellule, ce qui conduit à l'activation de mTOR et à la synthèse des protéines.(11)
Vous pouvez voir comment TAT fonctionne dans la figure ci-dessous:
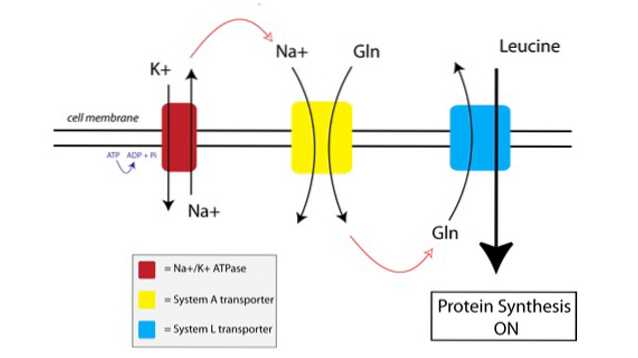
Tout d'abord, une pompe liée à la membrane appelée pompe ATPase sodium-potassium (Na + / K + ATPase, rouge dans la figure ci-dessus) utilise l'énergie de l'ATP pour déplacer le sodium à l'extérieur de la cellule, contre son gradient de concentration.
L'augmentation de la concentration de sodium à l'extérieur de la cellule est couplée à l'importation de glutamine par le transporteur du système A (jaune sur la figure). L'afflux de glutamine et de sodium dans la cellule attire également de l'eau supplémentaire, ce qui fait gonfler la cellule. Cela met la cellule dans un état anabolique, amorçant le mécanisme de synthèse des protéines pour l'activation.
Lorsque la glutamine s'accumule à des niveaux suffisamment élevés à l'intérieur de la cellule, les transporteurs du système L (en bleu sur la figure) sont activés, qui transportent la glutamine à l'extérieur de la cellule en échange de l'absorption de la leucine. L'entrée de la leucine dans la cellule est le déclencheur de la synthèse des protéines.
Bien que cela ait été une excellente leçon de biochimie jusqu'à présent, la découverte du TAT n'est pas seulement importante pour les biologistes cellulaires. Maintenant que nous savons comment le volume cellulaire est couplé au transport des acides aminés et à la synthèse des protéines, nous pouvons concevoir plusieurs stratégies nutritionnelles pour maximiser le processus quand cela compte - pendant la période critique après l'entraînement.
La synthèse des protéines dépend totalement de l'hydratation cellulaire - si vous êtes même un peu déshydraté, la capacité de récupérer après un entraînement intense est totalement compromise. Obtenir beaucoup d'eau est une évidence ici, mais l'eau seule ne suffit pas.
Les électrolytes tels que le sodium, le potassium, le chlorure et le phosphate fonctionnent également comme des «osmolytes» car ils attirent l'eau dans la cellule. Après une séance d'entraînement intense, nous avons besoin d'eau, d'acides aminés et d'électrolytes pour maximiser le processus de volumisation cellulaire qui entraîne la synthèse des protéines.
Le sodium, le magnésium, le calcium, le potassium, le phosphate et le chlorure (pour n'en nommer que quelques-uns) sont tous importants ici. À un niveau minimal (sauf si vous le faites sous l'avis de votre médecin), ne craignez pas le sodium avant ou après l'entraînement. Si vous êtes appauvri en sodium, la pompe de votre entraînement sera presque inexistante et le sodium est nécessaire à l'absorption de la glutamine.
Pour éliminer les conjectures, Surge® Workout Fuel et Plazma ™ sont conçus avec des rapports électrolytiques idéaux pour soutenir le volume cellulaire et la synthèse des protéines.
L'absorption de la glutamine dans la cellule provoque une volumisation cellulaire, amorçant les cellules musculaires pour la synthèse des protéines. Comme mentionné, un muscle plein / volumisé est un muscle anabolique. En plus de stimuler le transport des acides aminés, la volumisation cellulaire augmente également la synthèse du glycogène et inhibe la dégradation des protéines.(4-6)
La synthèse des protéines est supprimée par l'épuisement de la glutamine, ce qui a d'énormes implications pour les athlètes qui s'entraînent intensément. Après une séance d'entraînement intense, une réponse inflammatoire est montée, ce qui permet aux cellules immunitaires de pénétrer dans le tissu musculaire écrasé pour commencer le processus de réparation / reconstruction.(14)
La glutamine est si rapidement absorbée par les cellules immunitaires qu'elle est considérée comme le «carburant du système immunitaire.»(15) Sans surprise, il a été démontré qu'un entraînement intense provoque une déplétion plasmatique de la glutamine.(16-18)
Pour cette raison, les besoins en glutamine augmentent dans la période post-entraînement, où la réponse immunitaire locale peut être en compétition pour la disponibilité de la glutamine pour amorcer les cellules musculaires pour l'absorption d'acides aminés et la synthèse des protéines.
Pré-chargement les cellules contenant de la glutamine peuvent également potentiellement réduire le «temps de latence» associé à l'activation de la synthèse protéique par la leucine. Si vous ne le faites pas déjà, prenez 10 à 15 g de glutamine ou de peptides de glutamine immédiatement après l'entraînement. Parce que les BCAA sont un autre substrat préféré pour la synthèse de la glutamine musculaire et qu'il a également été démontré qu'ils augmentent la production de glutamine musculaire (19-21), les BCAA et la leucine sont également utiles pendant la période pré-entraînement pour aider à maximiser la production de glutamine endogène.
Note de l'éditeur: L'ensemble du protocole Plamza / MAG-10, avec sa nutrition pré-entraînement, entraînement et post-entraînement, répond aux recommandations de l'auteur pour charger les muscles avec des BCAA et de la leucine.
Récemment, il a été découvert que la consommation d'EAA augmente l'expression des transporteurs d'acides aminés du système A et du système L.(9) Il est important de noter que cela se produit au «niveau post-transcriptionnel», ce qui signifie au niveau de la synthèse des protéines, où les ARNm existants sont traduits en protéines.
Comparez cela avec "Denovo»Expression de protéines - où la synthèse, le traitement et le transport de nouveaux ARNm peuvent prendre 16 heures ou plus - l'activation post-transcriptionnelle de la synthèse des protéines peut se produire en quelques minutes à quelques heures, permettant aux cellules d'augmenter rapidement le niveau de protéines particulières en cas de besoin.
Soudainement, nous sommes plus incités à mettre en place un solide plan de nutrition péri-entraînement - l'apport d'EAA pendant les périodes pré et péri-entraînement est très rentable après l'entraînement en augmentant l'expression des transporteurs d'acides aminés, amorçant la cellule pour une absorption maximale d'acides aminés. et activation de la synthèse des protéines.
Les isolats ou hydrolysats de protéines à absorption rapide tels que Mag-10® Protein Pulsing Protocol ™ ou Plazma ™ pendant les périodes pré et péri-entraînement sont idéaux ici.
L'insuline est l'hormone la plus anabolique du corps. En plus d'activer directement la synthèse des protéines, l'insuline augmente également la translocation des transporteurs d'acides aminés du système A vers la membrane cellulaire.(22)
Cela signifie que l'insuline provoque l'affichage de plus de transporteurs du système A sur la membrane cellulaire, prêts à conduire plus de glutamine dans la cellule. Plus de glutamine conduit à plus de volume cellulaire, ce qui entraîne plus de leucine dans la cellule, conduisant finalement à une plus grande synthèse des protéines.
Alors que les EAA augmentent le expression des transporteurs AA, c'est le signal de l'insuline qui leur permet d'être affichés à la surface de la cellule, prêts à transporter de nouveaux acides aminés dans la cellule.
C'est encore une autre raison pour laquelle les glucides avant et pendant l'entraînement sont une bonne idée, sauf si vous êtes en mode de perte de graisse extrême: l'insuline augmente la capacité de transport cellulaire des acides aminés.
Les glucides augmentent les niveaux d'insuline, mais certains acides aminés peuvent également être utilisés pour potentialiser la libération d'insuline. La glutamine est un puissant activateur des hormones «incrétines», qui rendent les cellules productrices d'insuline dans le pancréas plus sensibles au glucose.(23) La glycine potentialise également la libération d'insuline par un mécanisme différent.
Bien que les glucides post-entraînement seuls augmentent les niveaux d'insuline, la combinaison de ces acides aminés potentialisant l'insuline avec des glucides surchargera votre pancréas pour encore plus de libération d'insuline. Bien qu'il soit bon de maintenir les niveaux d'insuline sur le côté inférieur la plupart du temps, l'augmentation des niveaux d'insuline pendant la période péri-entraînement maximise le transport des acides aminés, le volume cellulaire et la synthèse des protéines tout en supprimant la dégradation des protéines.
Le type d'entraînement anaérobie intense nécessaire pour construire des muscles de qualité entraîne une production considérable de lactate, ce qui diminue le pH musculaire. Cela entraîne une fatigue et une faiblesse musculaires précoces, mais certains transporteurs d'acides aminés, y compris le système A, sont également inhibés par un pH bas.(13)
Lorsque le pH musculaire est bas, l'absorption d'acides aminés est réduite, ce qui supprime l'activation de la synthèse protéique par mTOR.(24) Il a également été démontré qu'une faible inhibition du pH des transporteurs d'acides aminés du système A augmente la dégradation des protéines.(25)
C'est là qu'intervient la bêta-alanine. L'augmentation des niveaux de carnosine musculaire agit comme un tampon acide naturel, prolongeant le seuil anaérobie en limitant la diminution du pH musculaire due à l'entraînement.
La bêta-alanine a une autre fonction importante: aider à maintenir la synthèse des protéines et à la mettre en ligne plus rapidement après un entraînement intense en empêchant l'atténuation du transport des acides aminés.
Pour augmenter les niveaux de carnosine dans les muscles, prenez deux comprimés de Beta-7 ™ trois fois par jour.
Lors des séances d'entraînement intenses, la synthèse des protéines est réduite et la dégradation des protéines est activée. Ceci est inévitable pour tout athlète d'entraînement dur. Cependant, la mesure dans laquelle nous pouvons minimiser les effets cataboliques de l'entraînement et le retour rapide au «mode anabolique» pendant la période post-entraînement détermine en fin de compte l'efficacité avec laquelle nous récupérerons et grandirons.
Le timing des macronutriments est important, mais c'est un moyen pour arriver à une fin. Le volume cellulaire est le principal moteur du transport des acides aminés et de la synthèse des protéines. En comprenant comment le transport des acides aminés se produit et comment il est régulé par le volume cellulaire, nous pouvons obtenir plus de leucine dans les cellules musculaires détruites plus rapidement, alimentant ainsi le feu anabolique et conduisant finalement à de meilleurs gains.
Les stratégies ci-dessus sont efficaces, pratiques et basées sur les dernières recherches scientifiques. Utilisez-les comme modèle pour faire passer votre progression d'entraînement au niveau supérieur.
J'attends avec impatience vos questions sur le LiveSpill!



Personne n'a encore commenté ce post.